
酵母双杂交实验流程.doc

听容****55


1/9

2/9

3/9

4/9
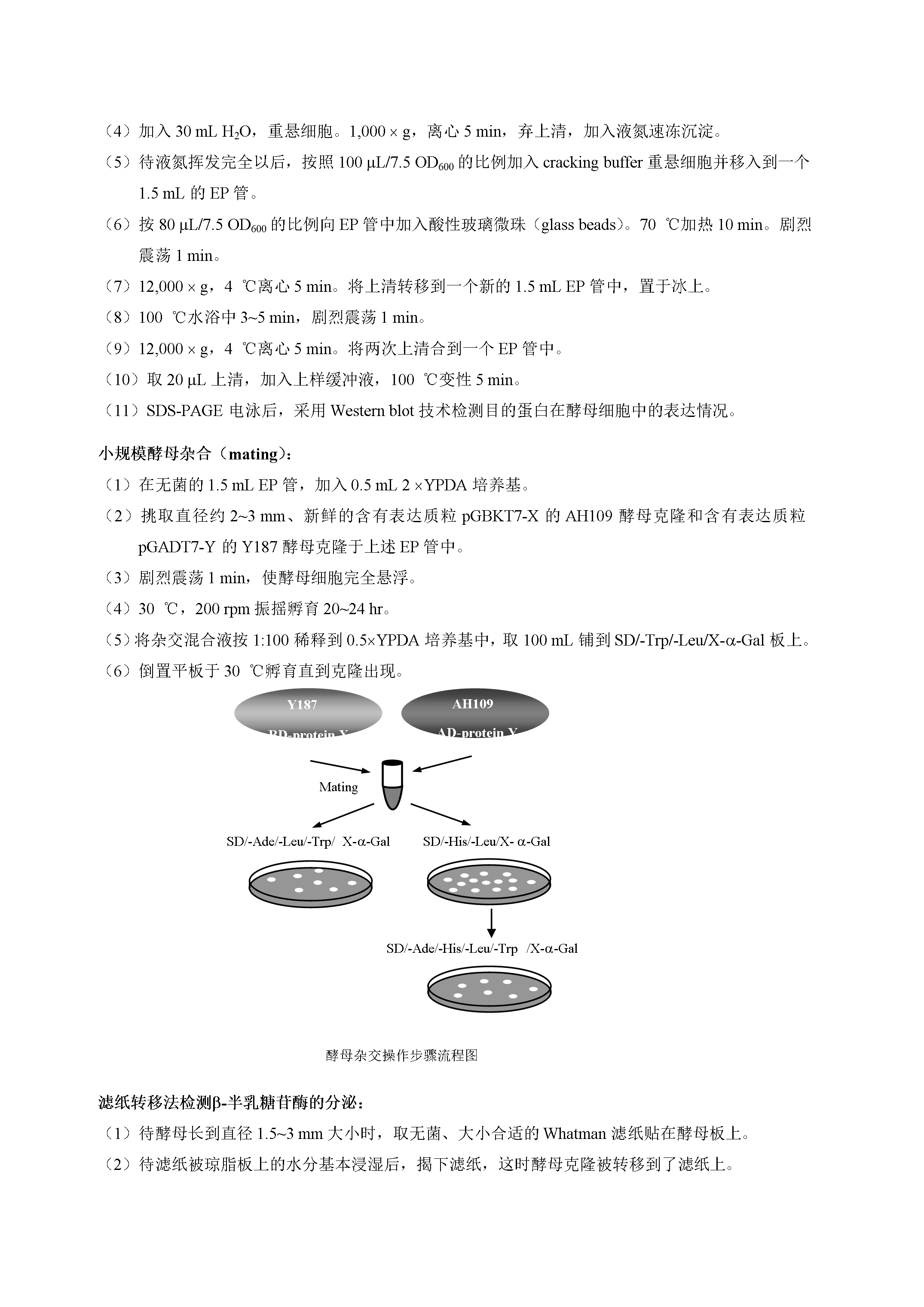
5/9
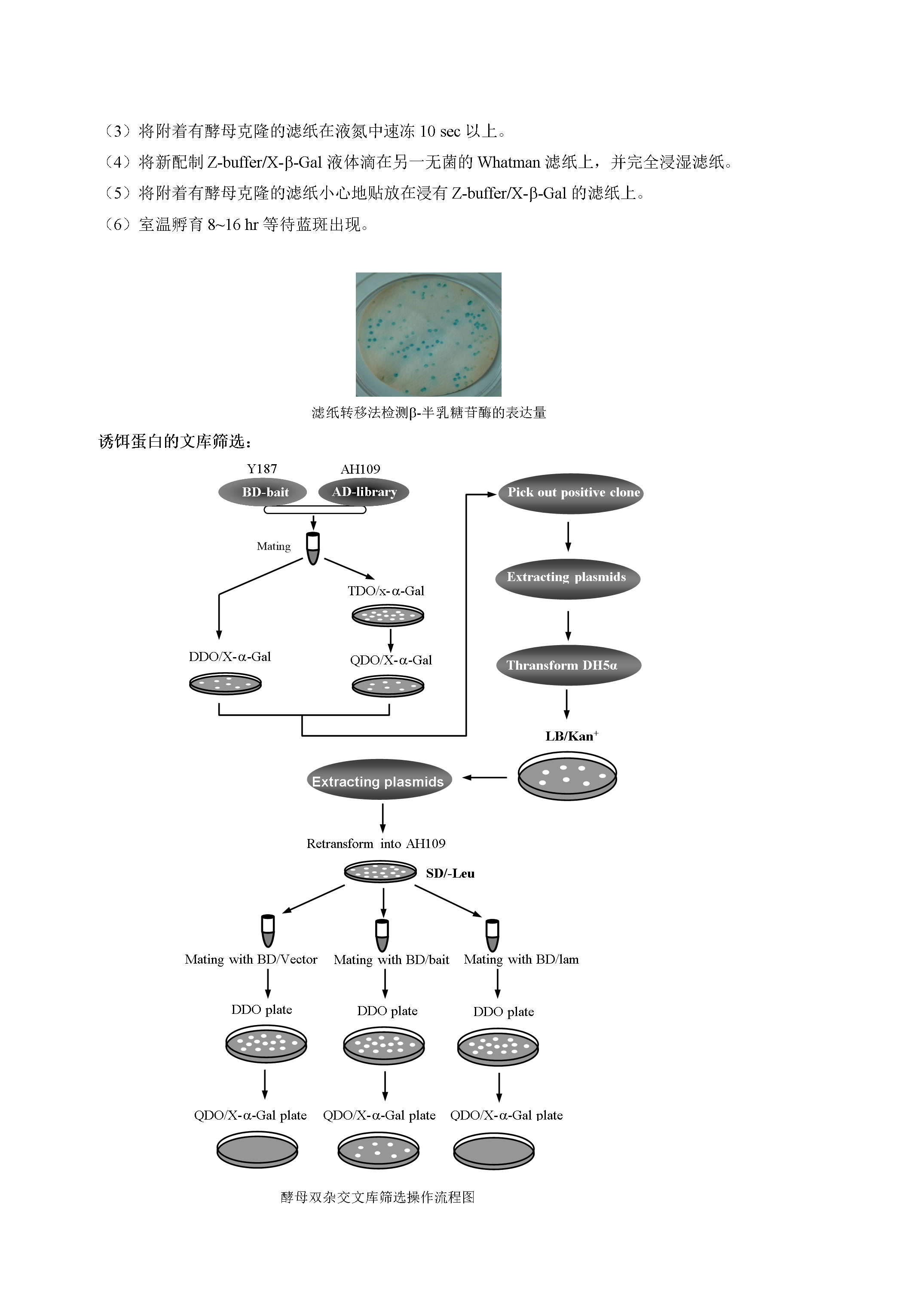
6/9

7/9

8/9

9/9
在线预览结束,喜欢就下载吧,查找使用更方便
相关资料

酵母双杂交实验流程.pdf
模块七蛋白质之间的相互作用1.实验目的本实验以重组质粒和酵母细胞为材料,学习检测蛋白质相互作用的基本原理和技术方法。主要介绍酵母双杂交的基本原理与操作技术;让学生了解和掌握酵母双杂交系统的应用;掌握酵母感受态的制备的基本原理和主要的操作步骤。2.实验原理1989年Fields和Song等人根据当时人们对真核生物转录起始过程调控的认识(即细胞内基因转录的起始需要转录激活因子的参与),提出并建立了酵母双杂交系统。该系统作为发现和研究活细胞体内的蛋白质与蛋白质之间的相互作用的技术平台,近几年得到了广泛的运用和发

酵母双杂交实验流程.doc
模块七蛋白质之间的相互作用1.实验目的本实验以重组质粒和酵母细胞为材料学习检测蛋白质相互作用的基本原理和技术方法。主要介绍酵母双杂交的基本原理与操作技术;让学生了解和掌握酵母双杂交系统的应用;掌握酵母感受态的制备的基本原理和主要的操作步骤。2.实验原理1989年Fields和Song等人根据当时人们对真核生物转录起始过程调控的认识(即细胞内基因转录的起始需要转录激活因子的参与)提出并建立了酵母双杂交系统。该系统作为发现和研究活细胞体内的蛋白质与蛋白质之间的相互作用的技术平台近几年得到了广泛的运用
酵母双杂交实验流程.doc
模块七蛋白质之间的相互作用1.实验目的本实验以重组质粒和酵母细胞为材料学习检测蛋白质相互作用的基本原理和技术方法。主要介绍酵母双杂交的基本原理与操作技术;让学生了解和掌握酵母双杂交系统的应用;掌握酵母感受态的制备的基本原理和主要的操作步骤。2.实验原理1989年Fields和Song等人根据当时人们对真核生物转录起始过程调控的认识(即细胞内基因转录的起始需要转录激活因子的参与)提出并建立了酵母双杂交系统。该系统作为发现和研究活细胞体内的蛋白质与蛋白质之间的相互作用的技术平台近几年得到了广泛的运用
酵母双杂交实验流程.doc
模块七蛋白质之间的相互作用1.实验目的本实验以重组质粒和酵母细胞为材料学习检测蛋白质相互作用的基本原理和技术方法。主要介绍酵母双杂交的基本原理与操作技术;让学生了解和掌握酵母双杂交系统的应用;掌握酵母感受态的制备的基本原理和主要的操作步骤。2.实验原理1989年Fields和Song等人根据当时人们对真核生物转录起始过程调控的认识(即细胞内基因转录的起始需要转录激活因子的参与)提出并建立了酵母双杂交系统。该系统作为发现和研究活细胞体内的蛋白质与蛋白质之间的相互作用的技术平台近几年得到了广泛的运用

酵母双杂交实验流程(精).doc
(完整word)酵母双杂交实验流程(精)(完整word)酵母双杂交实验流程(精)(完整word)酵母双杂交实验流程(精)模块七蛋白质之间的相互作用1.实验目的本实验以重组质粒和酵母细胞为材料,学习检测蛋白质相互作用的基本原理和技术方法。主要介绍酵母双杂交的基本原理与操作技术;让学生了解和掌握酵母双杂交系统的应用;掌握酵母感受态的制备的基本原理和主要的操作步骤。2.实验原理1989年Fields和Song等人根据当时人们对真核生物转录起始过程调控的认识(即细胞内基因转录的起始需要转录激活因子的参与,提出并建