
假基因简介0.ppt

胜利****实阿


1/10

2/10

3/10
4/10

5/10
6/10

7/10

8/10

9/10

10/10
亲,该文档总共20页,到这已经超出免费预览范围,如果喜欢就直接下载吧~
相关资料

假基因简介0.ppt
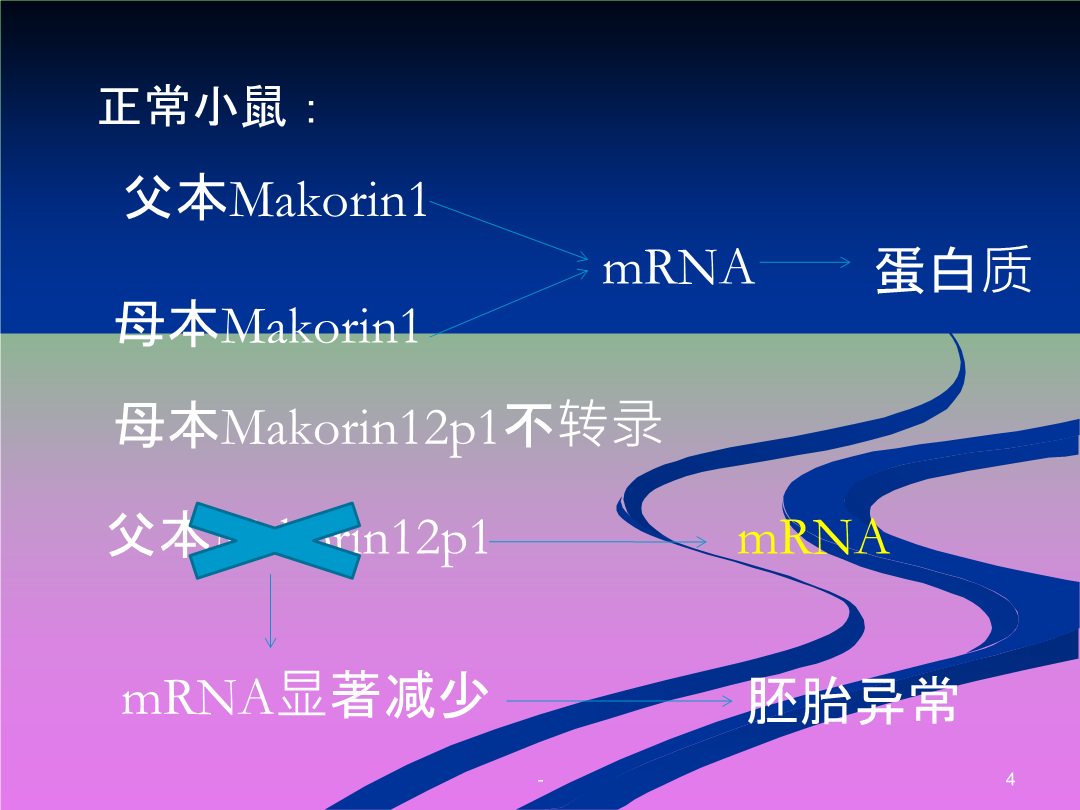
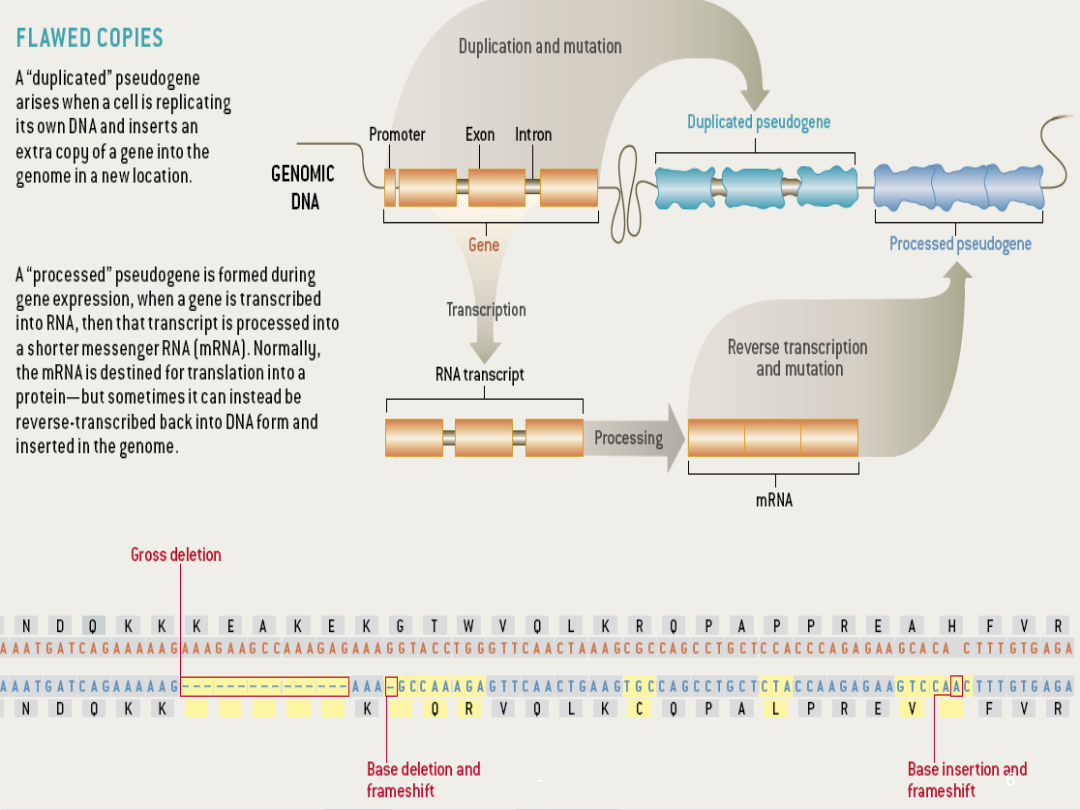
假基因的发现定义父本Makorin1分类加工假基因起源于反转录转座作用,即通过DNA转录为mRNA后,再由mRNA反转录成cDNA,然后随机整合到基因组DNA中而产生。因此它们也称为返座假基因或逆转座型假基因1.两末端都有很短的定向重复序列2.3’末端带有多聚腺嘌呤尾部序列3.编码区密码子的突变和终止密码子的提前出现4.缺乏内含子和启动子区域5.目前只在真核生物中发现加工假基因,且其形成机制多与RNA聚合酶Ⅱ有关1.常位于其起源功能基因附近2.结构与同源功能基因类似3.仍然含有内含子4.目前发现细菌和真核

假基因简介PPT0.ppt
假基因的发现定义父本Makorin1分类加工假基因起源于反转录转座作用,即通过DNA转录为mRNA后,再由mRNA反转录成cDNA,然后随机整合到基因组DNA中而产生。因此它们也称为返座假基因或逆转座型假基因1.两末端都有很短的定向重复序列2.3’末端带有多聚腺嘌呤尾部序列3.编码区密码子的突变和终止密码子的提前出现4.缺乏内含子和启动子区域5.目前只在真核生物中发现加工假基因,且其形成机制多与RNA聚合酶Ⅱ有关1.常位于其起源功能基因附近2.结构与同源功能基因类似3.仍然含有内含子4.目前发现细菌和真核

假基因简介.ppt
假基因的发现定义父本Makorin1分类加工假基因起源于反转录转座作用,即通过DNA转录为mRNA后,再由mRNA反转录成cDNA,然后随机整合到基因组DNA中而产生。因此它们也称为返座假基因或逆转座型假基因1.两末端都有很短的定向重复序列2.3’末端带有多聚腺嘌呤尾部序列3.编码区密码子的突变和终止密码子的提前出现4.缺乏内含子和启动子区域5.目前只在真核生物中发现加工假基因,且其形成机制多与RNA聚合酶Ⅱ有关1.常位于其起源功能基因附近2.结构与同源功能基因类似3.仍然含有内含子4.目前发现细菌和真核

假基因简介.ppt
假基因的发现定义父本Makorin1分类加工假基因起源于反转录转座作用,即通过DNA转录为mRNA后,再由mRNA反转录成cDNA,然后随机整合到基因组DNA中而产生。因此它们也称为返座假基因或逆转座型假基因1.两末端都有很短的定向重复序列2.3’末端带有多聚腺嘌呤尾部序列3.编码区密码子的突变和终止密码子的提前出现4.缺乏内含子和启动子区域5.目前只在真核生物中发现加工假基因,且其形成机制多与RNA聚合酶Ⅱ有关1.常位于其起源功能基因附近2.结构与同源功能基因类似3.仍然含有内含子4.目前发现细菌和真核

假基因简介PPT.ppt
假基因的发现定义父本Makorin1分类加工假基因起源于反转录转座作用,即通过DNA转录为mRNA后,再由mRNA反转录成cDNA,然后随机整合到基因组DNA中而产生。因此它们也称为返座假基因或逆转座型假基因1.两末端都有很短的定向重复序列2.3’末端带有多聚腺嘌呤尾部序列3.编码区密码子的突变和终止密码子的提前出现4.缺乏内含子和启动子区域5.目前只在真核生物中发现加工假基因,且其形成机制多与RNA聚合酶Ⅱ有关1.常位于其起源功能基因附近2.结构与同源功能基因类似3.仍然含有内含子4.目前发现细菌和真核