
基因组进化的模式ppt课件.ppt

lj****88


1/10
2/10
3/10

4/10

5/10

6/10

7/10

8/10

9/10

10/10
亲,该文档总共52页,到这已经超出免费预览范围,如果喜欢就直接下载吧~
相关资料

基因组进化的模式ppt课件.ppt
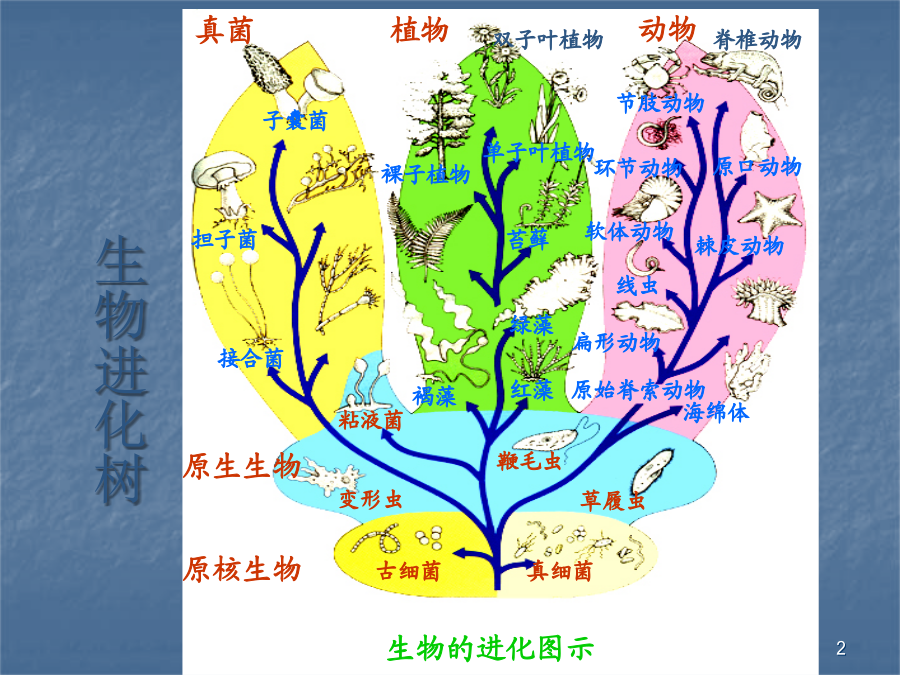
第八章基因组进化的模式生物进化树分子水平生命史的三大阶段原始气体冷凝汇流成海洋含生命化石的最古老的岩石一、遗传系统的起源蛋白质起源说奥巴林的团聚体(coacervate)假说(1924):用蛋白质(白明胶)和多糖(阿拉伯胶)混合得到团聚体福克斯用类蛋白质加热得到微球体S.Miller(1953)的模拟实验澳大利亚陨石中发现(1959)氨基酸、嘧啶和脂肪酸用电击提供能量,从简单小分子可得到复杂大分子。生物分子形成的传统理论核酸起源说1.1RNA世界假说TheNobelPrizeinChemistry1989

基因组进化模式ppt课件.ppt
XiaofangXieCollegeofLifeScienceFujianAgricultureandForestryUniversityE-mail:xxf317@yahoo.com.cn基因组的进化模式基因组进化生命起源COLLEGEOFLIFESCIENCE“生命的起源自然发生说”成立的基本条件最早的生化系统什么是生命?最早的生命形式是什么?勾勒出与现有生命系统相似的生物大分子合成的地球化学过程,生物大分子是如何从随机组合到有序组装,显示部分生命相关的生化特性的遗传/生命起源Q1:蛋白质?----不

基因组进化的分子基础PPT课件.ppt
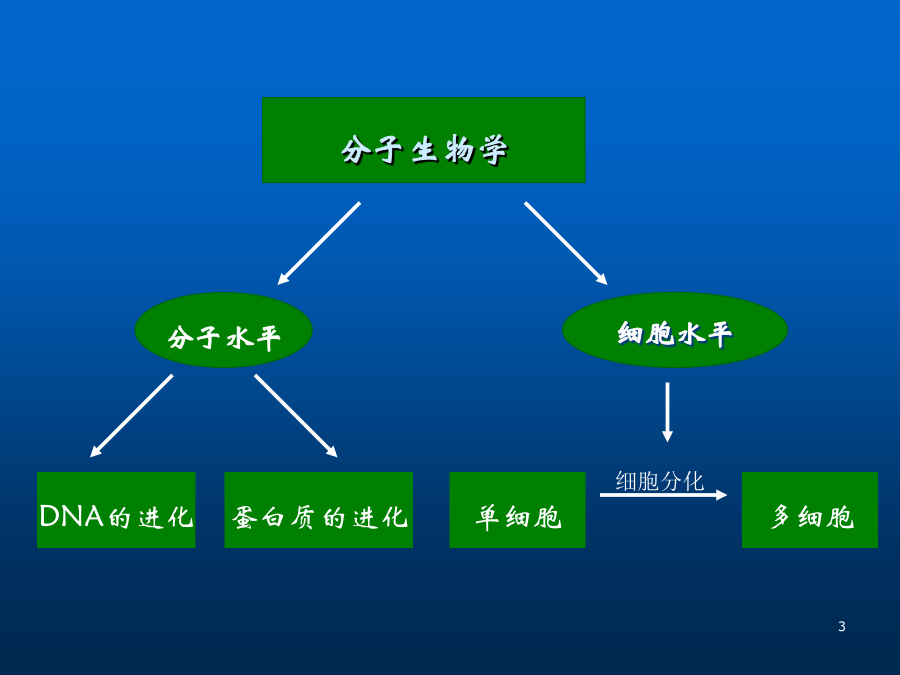
基因组进化的分子基础基因组进化的分子基础:突变重组转座突变1.1突变的机制♦自发性损伤(复制中的损伤、碱基的自发性化学改变、自发脱碱基、细胞的代谢产物对DNA的损伤)♦物理因素引起的损伤(电离辐射、紫外线、热诱变等)♦化学因素引起的损伤(烷化剂、碱基类似物、嵌入试剂等)1.1.1自发性损伤错配突变纯化学的碱基配对差错率为:5%~10%为维持基因组的稳定性,DNA的复制必须增加几个数量级,提高DNA复制的精确性有2种方法:♪渗入碱基的筛选♪错配碱基的校正碱基异构式引起DNA复制过程的错误-----自发突变碱

比较基因组与分子进化ppt课件.ppt
吴坤陆分子进化除RNA病毒外,几乎所有生命的蓝图都用DNA书写,进化的第一原因是基因突变。Humanfishβ-globin分子进化的速率约为10-9,每年每个氨基酸座位的1×10-9进化速率为分子进化速率的单位,即1×10-9为1鲍林。1.分子进化速率具有恒定性对于各种生物物种的每一个蛋白质,用每一个位点每年发生的氨基酸替换的次数为标准衡量分子进化的速率是大致恒定的,只要该分子的功能和三维结构保持不变;2.分子进化的保守性不同对生物生存制约性大的生物大分子进化速度慢。生物大分子内部功能区结构变化的速度较

水稻基因组的进化选择模式研究.docx
水稻基因组的进化选择模式研究水稻基因组的进化选择模式研究水稻(Oryzasativa)是人类最主要的粮食作物之一,在全球范围内有着广泛的种植和消费。水稻的基因组组成复杂,包括数万个基因以及大量的非编码RNA序列、编码蛋白序列和等位基因等元素。这些基因元素在水稻生长发育、抗性和适应性方面发挥着重要的作用。在水稻基因组中,不同区域的基因组组成和遗传学特征存在巨大差异。这些差异的形成与环境压力、群体遗传力学和进化选择等因素密切相关。本文将综述水稻基因组的进化选择特点,重点分析水稻中与适应性和抗性密切相关的基因组