
植物生理学信号转导.1PPT课件.ppt

ca****ng


1/10
2/10
3/10
4/10

5/10

6/10
7/10
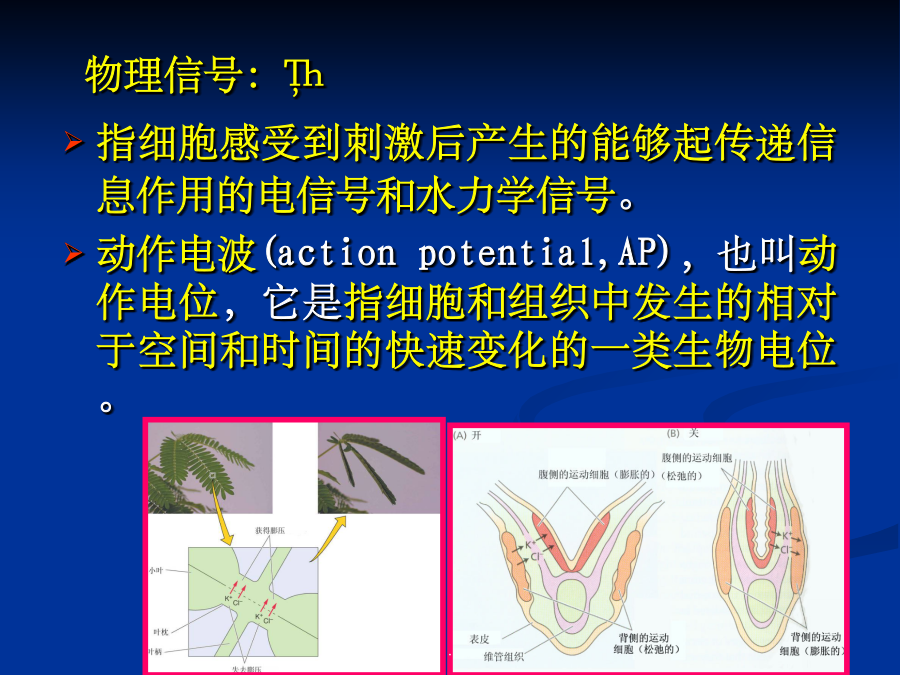
8/10

9/10
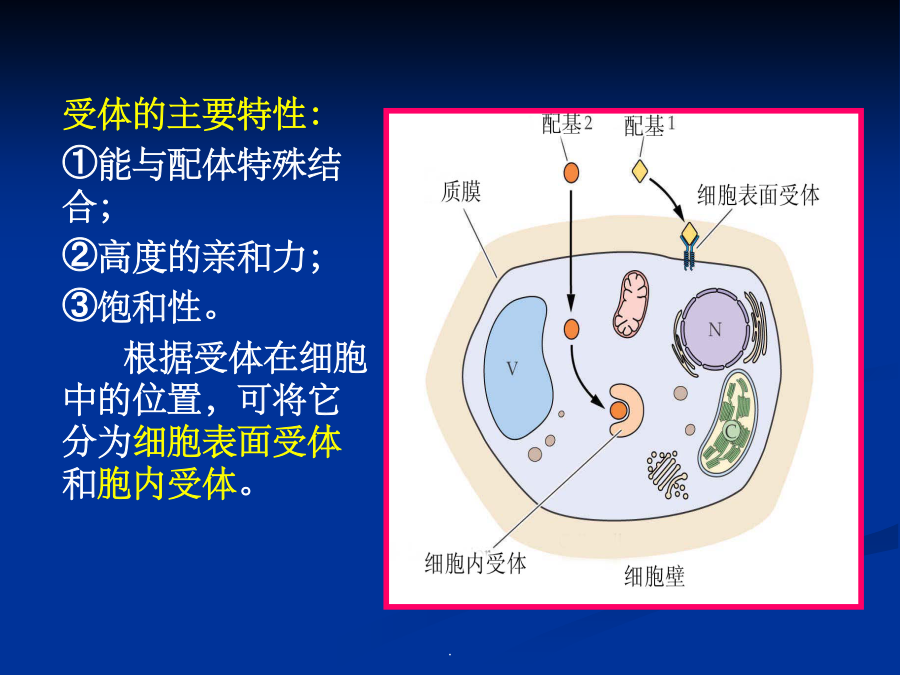
10/10
亲,该文档总共44页,到这已经超出免费预览范围,如果喜欢就直接下载吧~
相关资料

植物生理学信号转导.1PPT课件.ppt
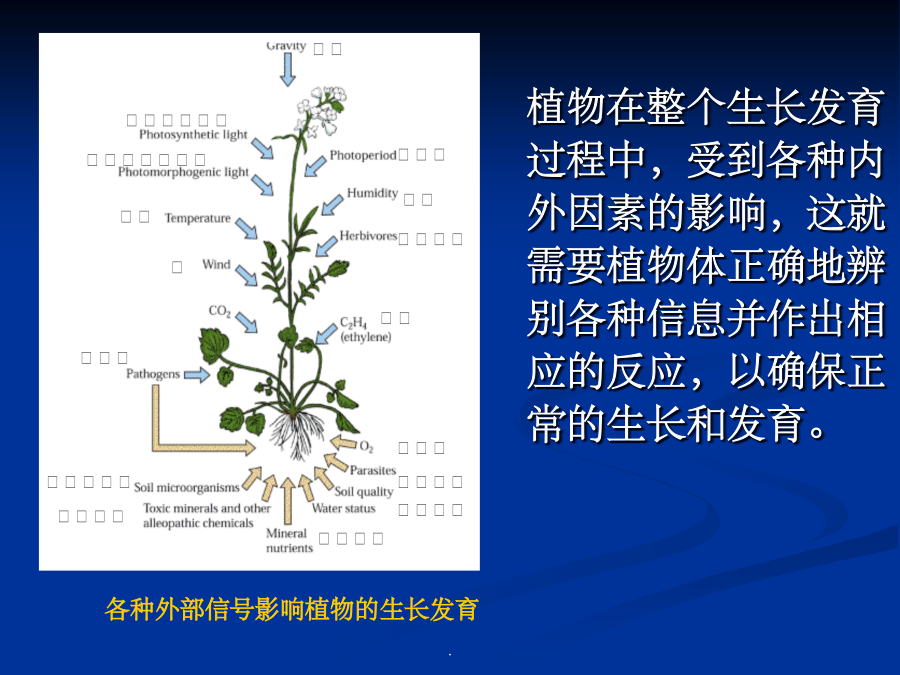
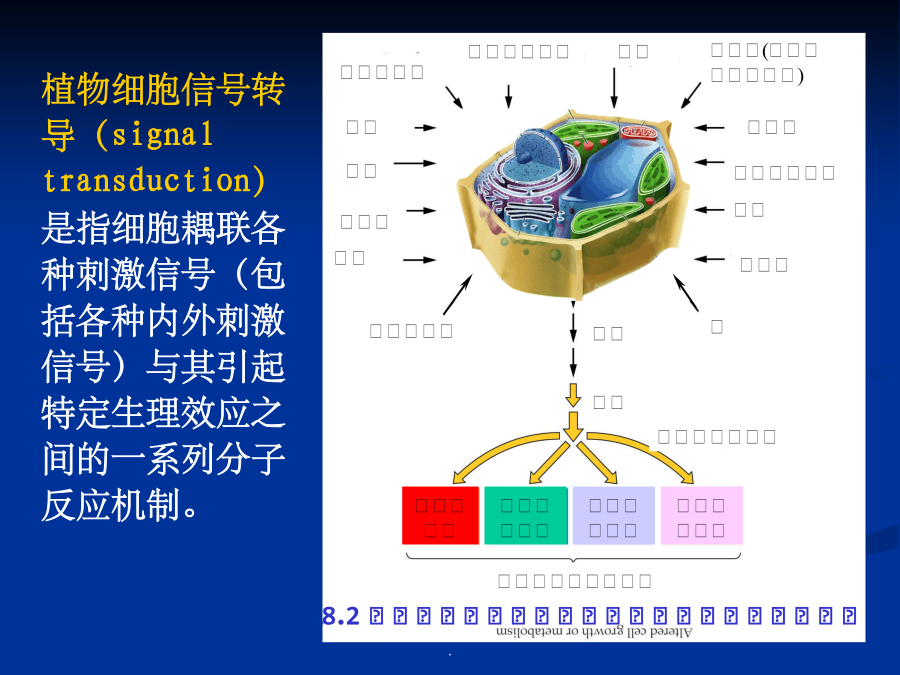
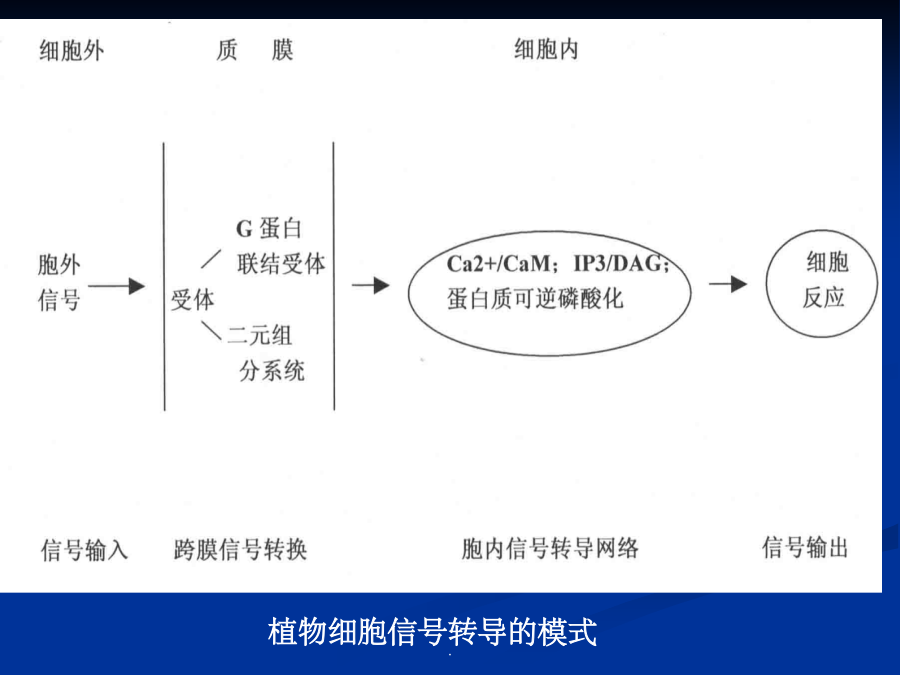
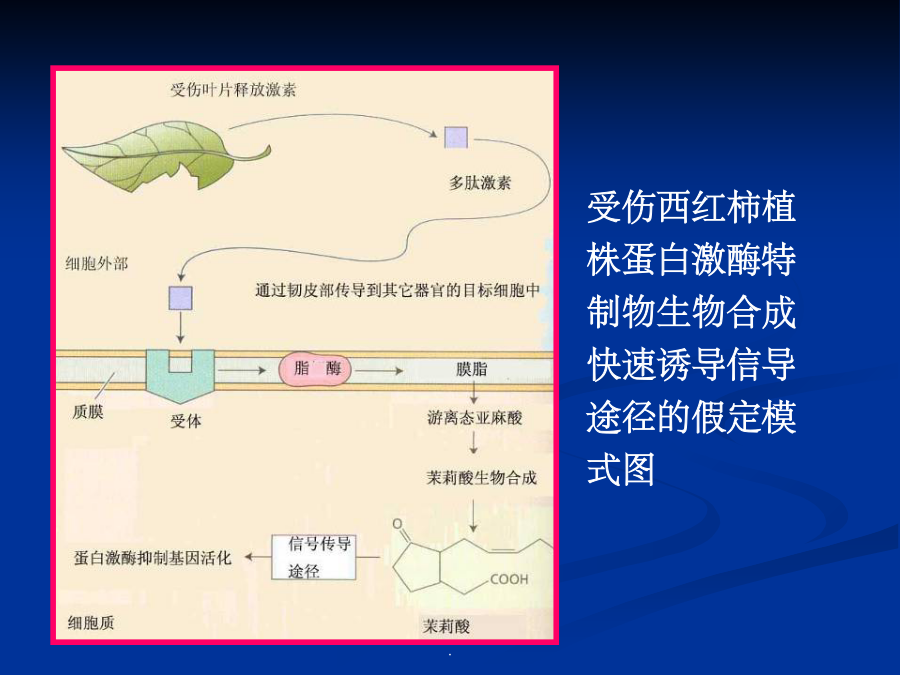
第七章细胞信号转导各种外部信号影响植物的生长发育植物细胞信号转导(signaltransduction)是指细胞耦联各种刺激信号(包括各种内外刺激信号)与其引起特定生理效应之间的一系列分子反应机制。植物细胞信号转导的模式化学信号(chemicalsignals):细胞感受刺激后合成并传递到作用部位引起生理反应的化学物质。激素、病原因子等,化学信号也叫做配体(ligand)当植物的一张叶片被虫咬伤后,会诱导本叶和其它叶产生蛋白酶抑制物(PIs)等,以阻碍病原菌或害虫进一步侵害。如果伤害后立即除去受害叶,则其

植物生理学1PPT课件.ppt
涝害生理与植物抗涝性涝害生理与植物抗涝性涝害的定义:水分过多对植物的伤害称为涝害涝害是因降水过多、土壤含水量过大,使作物生长受到损害的现象,是田间水分过多,出现“渍”、“淹”、“涝”,致使作物生长受到危害的现象。涝害对植物的伤害植物抗涝性的生理基础提高植物抗涝性的途径

植物生理学细胞及信号转导PPT课件.ppt
根据细胞的进化程度,可将其分为两大类型:原核细胞(prokaryoticcell)和真核细胞(eukaryoticcell)。细菌和蓝藻是原核细胞的典型代表,此外支原体、衣原体、立克次体、放线菌等也都是原核细胞。原核细胞一般体积很小,直径为0.2~10μm不等,没有典型的细胞核,即没有核膜将它的遗传物质与细胞质分开,只有一个由裸露的环状DNA分子构成的拟核体(nucleoid)。除核糖体、类囊体外,一般不存在其它细胞器,原核细胞以无丝分裂(amitosis)方式进行繁殖。由真核细胞构成的有机体称为真核生物

植物生理学信号转导..ppt
第七章细胞信号转导各种外部信号影响植物的生长发育植物细胞信号转导(signaltransduction)是指细胞耦联各种刺激信号(包括各种内外刺激信号)与其引起特定生理效应之间的一系列分子反应机制。植物细胞信号转导的模式化学信号(chemicalsignals):细胞感受刺激后合成并传递到作用部位引起生理反应的化学物质。激素、病原因子等,化学信号也叫做配体(ligand)当植物的一张叶片被虫咬伤后,会诱导本叶和其它叶产生蛋白酶抑制物(PIs)等,以阻碍病原菌或害虫进一步侵害。如果伤害后立即除去受害叶,则其

植物生理学信号转导.ppt
第七章细胞信号转导各种外部信号影响植物的生长发育植物细胞信号转导(signaltransduction)是指细胞耦联各种刺激信号(包括各种内外刺激信号)与其引起特定生理效应之间的一系列分子反应机制。植物细胞信号转导的模式化学信号(chemicalsignals):细胞感受刺激后合成并传递到作用部位引起生理反应的化学物质。激素、病原因子等,化学信号也叫做配体(ligand)当植物的一张叶片被虫咬伤后,会诱导本叶和其它叶产生蛋白酶抑制物(PIs)等,以阻碍病原菌或害虫进一步侵害。如果伤害后立即除去受害叶,则其